3D基础模型助力植物表型:从分钟级到秒级的跨作物三维重建
The Turning Point of 3D Plant Phenotyping: 3D Foundation Models Enable Minute-to-Second Cross-Crop Reconstruction and Beyond
摘要 Abstract

三维植物表型(3D Plant Phenotyping)长期受限于流程复杂、吞吐量低等问题,主要原因在于多视角成像需求大、三维重建流水线脆弱、以及从重建几何到表型提取的额外成本。在低成本图像采集(如智能手机视频或稀疏多视角图像)条件下,这些限制被进一步放大。
本工作首次系统验证了3D基础模型(3DFMs)可以作为植物三维表型的前端几何基础,将传统COLMAP风格的分钟级初始化压缩到秒级。具体而言,作者提出了一个跨作物3D表型框架,核心创新包括:
- 3DFM前端替换:用VGGT和π³的前馈几何推理替代COLMAP的特征匹配+增量位姿估计
- 几何约束3DGS密集化:结合Mip-Splatting实现保边界的密集重建
- 少视角迭代增强:通过Difix3D+辅助的新视角合成补充稀疏观测
- 2D-to-3D语义迁移:将多视角2D分割结果反投影到3D空间实现器官实例分离
在26个植物序列上的实验表明,3DFM将平均前端初始化时间从6.52分钟压缩到1.58秒,同时保持可比的重建质量和器官级表型测量精度(叶面积估计比值0.95-1.06,叶倾角平均误差约2.04°)。
用3D基础模型替代COLMAP作为植物三维表型的前端初始化,将重建速度提升250倍,同时支持跨作物的器官级表型测量。
研究背景 Background
植物表型是连接基因型与环境的桥梁。相比2D成像,3D表型能更直接地捕获冠层架构、器官几何和植物结构,量化叶片朝向、光截获和建筑学特征。然而,高通量3D表型的核心挑战在于如何低成本、高效率地获取定量可用的3D植物结构。
传统3D植物表型经历了三个发展阶段:
- 主动传感阶段:LiDAR、结构光、ToF相机——精度高但成本高、部署复杂
- 被动多视角重建阶段:SfM/MVS——降低硬件门槛但依赖足够视角重叠,植物场景中重复纹理、自遮挡导致匹配失败
- 神经渲染阶段:NeRF/3DGS——提升表示能力但仍依赖COLMAP前端初始化
最近,3D基础模型(DUSt3R、MASt3R、VGGT、π³)的出现为突破前端瓶颈提供了新机会。这些模型将多视角几何推理从局部匹配+增量优化转变为基于学习先验的前馈式几何推理,但此前尚未在植物表型领域得到系统验证。
方法论 Methodology
框架整体设计为一个从低成本图像输入到可测量3D植物结构的连续数据变换链,包含四个核心阶段:
1. 3DFM快速初始化
用VGGT和π³作为前馈式初始化器,替代COLMAP的特征提取→匹配→增量位姿估计流程。3DFM直接从图像推断多视角几何,通过桥接转换步骤将模型特定输出标准化为统一的相机参数集和初始稀疏点云。π³采用无参考、置换等变设计,对输入顺序更具鲁棒性。
2. 几何约束3DGS密集化
采用Mip-Splatting作为密集化方法,其3D平滑和2D足迹控制在尺度变化下提升几何稳定性。通过3D协方差约束抑制退化或过度尖锐的基元,2D足迹约束减少投影中的锯齿和边界膨胀。在优化过程中,大梯度区域进行密集化以改善叶片边缘覆盖,低透明度基元被剪枝以抑制浮动伪影。
3. 少视角迭代增强
针对少视角(≤10个输入视角)场景,引入新视角合成→图像增强→输入更新→重重建的迭代方案。在现有视角间采样新视角位姿,用当前高斯表示渲染中间视图,再用Difix3D+进行残差渲染伪影修复。生成视图仅作为附加图像观测提供光度约束,不作为几何真值。
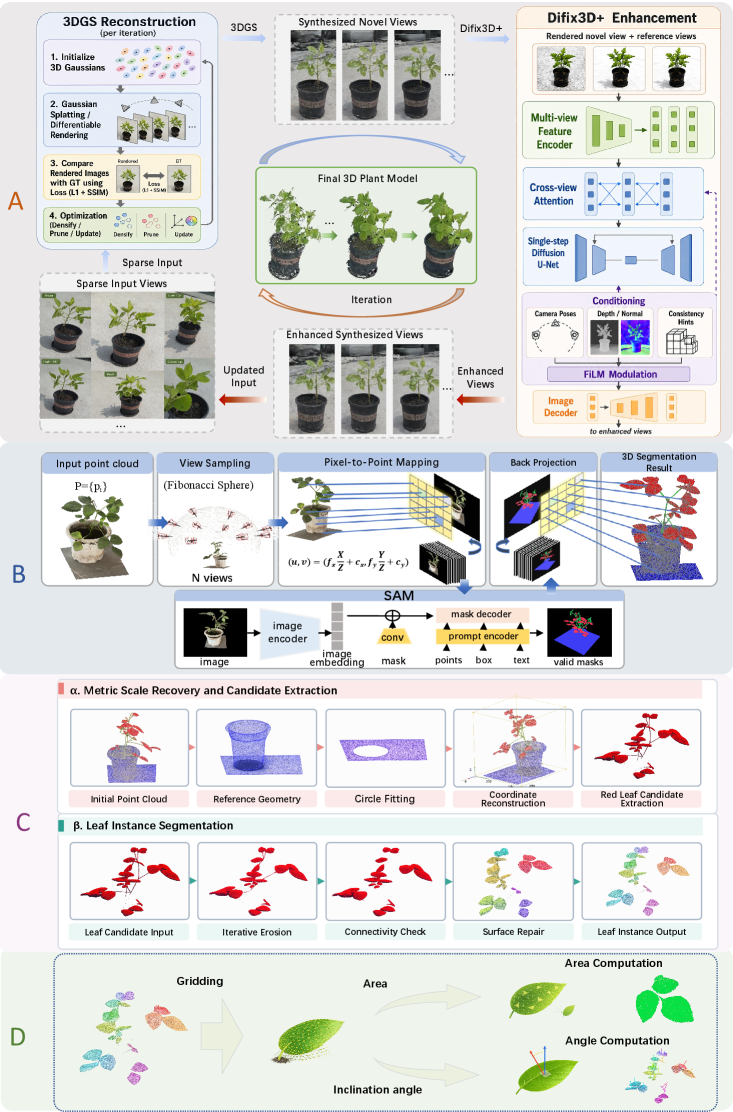
4. 2D-to-3D语义迁移与表型提取
采用投影式2D-to-3D语义迁移策略:多视角渲染→2D分割→像素-点对应建立→反投影→多视角融合。相比直接3D点云分割(PSegNet、PointTransformerV3等),该方法在叶片边界保持和遮挡鲁棒性方面显著更优。
尺度恢复利用花盆几何作为场景内度量参考,通过圆拟合估计重建花盆直径与实际直径的比值作为尺度因子。叶片实例分离采用几何驱动的层次策略:体素侵蚀→连通分析→边界恢复,将叶片点云分离为独立的叶片实例。最终通过Ball-Pivoting Algorithm重建开放网格,计算叶面积和叶倾角。
关键结果 Key Results
3DFM将前端初始化从分钟级压缩到秒级
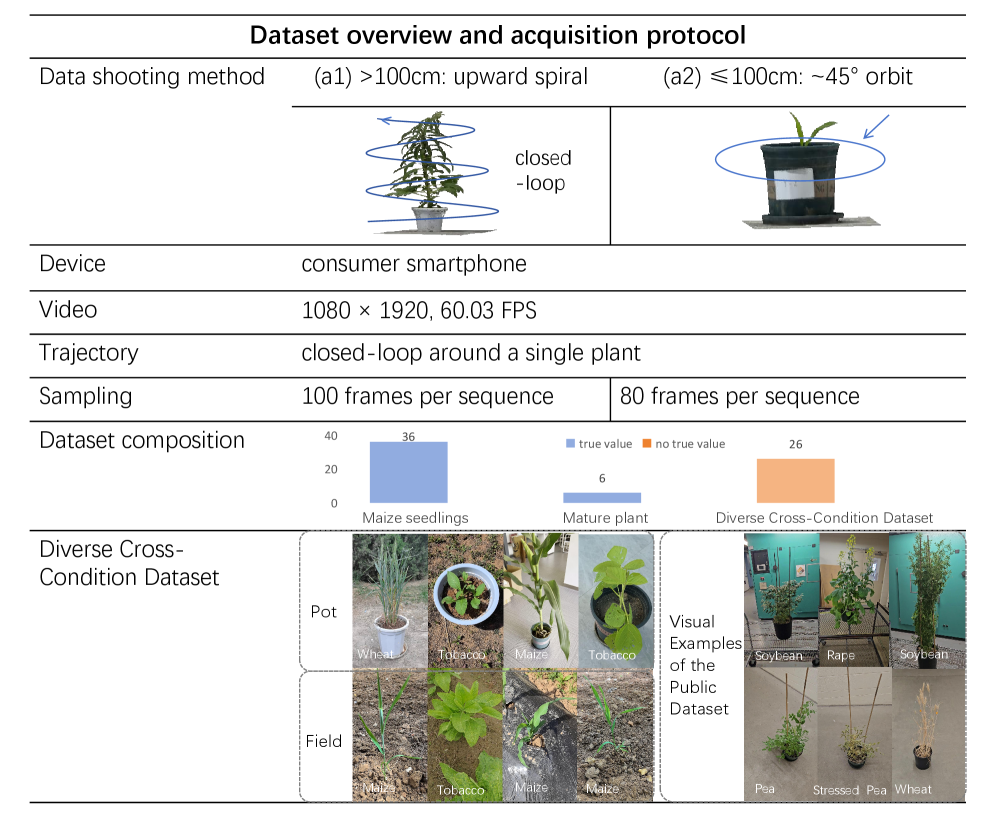
在26个植物序列(涵盖烟草、玉米、小麦、大豆、竹子、油菜、豆类、豌豆、西兰花等10种作物,室内/室外/田间三种环境)上的实验表明:
| 方法 | 平均前端时间 | SSIM | PSNR | LPIPS |
|---|---|---|---|---|
| COLMAP | 6.52 分钟 | 0.891 | 29.44 | 0.138 |
| VGGT | 1.55 秒 | 0.864 | 28.40 | 0.173 |
| π³ (本文) | 1.58 秒 | 0.886 | 29.27 | 0.144 |
π³在将初始化时间压缩250倍的同时,重建质量(SSIM/PSNR/LPIPS)与COLMAP非常接近。
少视角重建优势
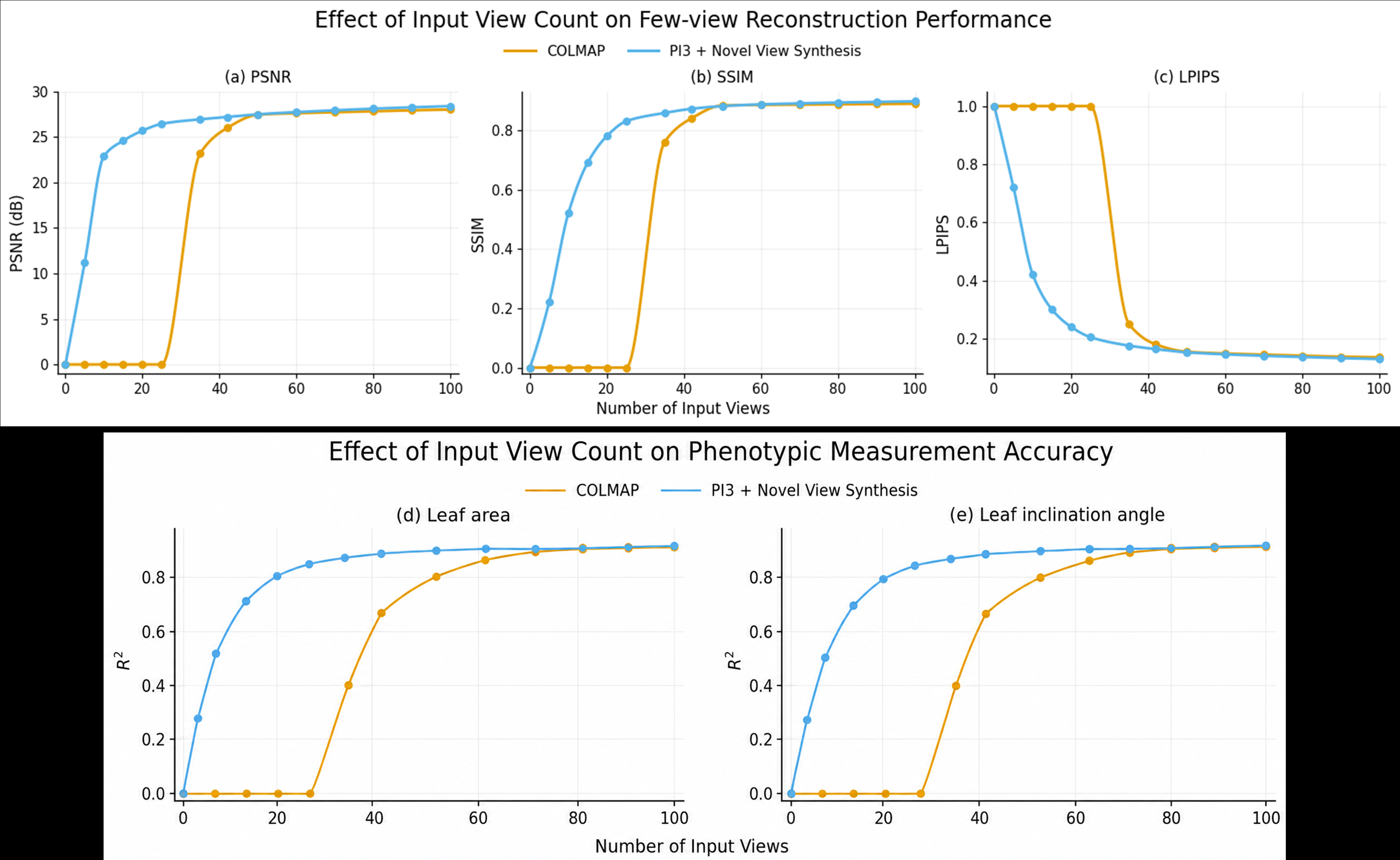
在低视角(≤20)范围内,COLMAP无法产生有效重建,而π³+新视角合成流水线在约10个视角时即达到可用水平。随着视角增加到50+,两者趋于收敛,表明3DFM的核心价值在于降低可用重建阈值,而非提升密集视角下的性能上限。
2D-to-3D语义迁移大幅优于直接3D分割
| 方法 | 大豆 OA | 大豆 mIoU | 玉米 OA | 玉米 mIoU |
|---|---|---|---|---|
| PSegNet | 0.703 | 0.466 | 0.781 | 0.396 |
| PointTransformerV3 | 0.768 | 0.517 | 0.764 | 0.431 |
| 本文方法 | 0.966 | 0.919 | 0.936 | 0.822 |
器官级表型测量精度
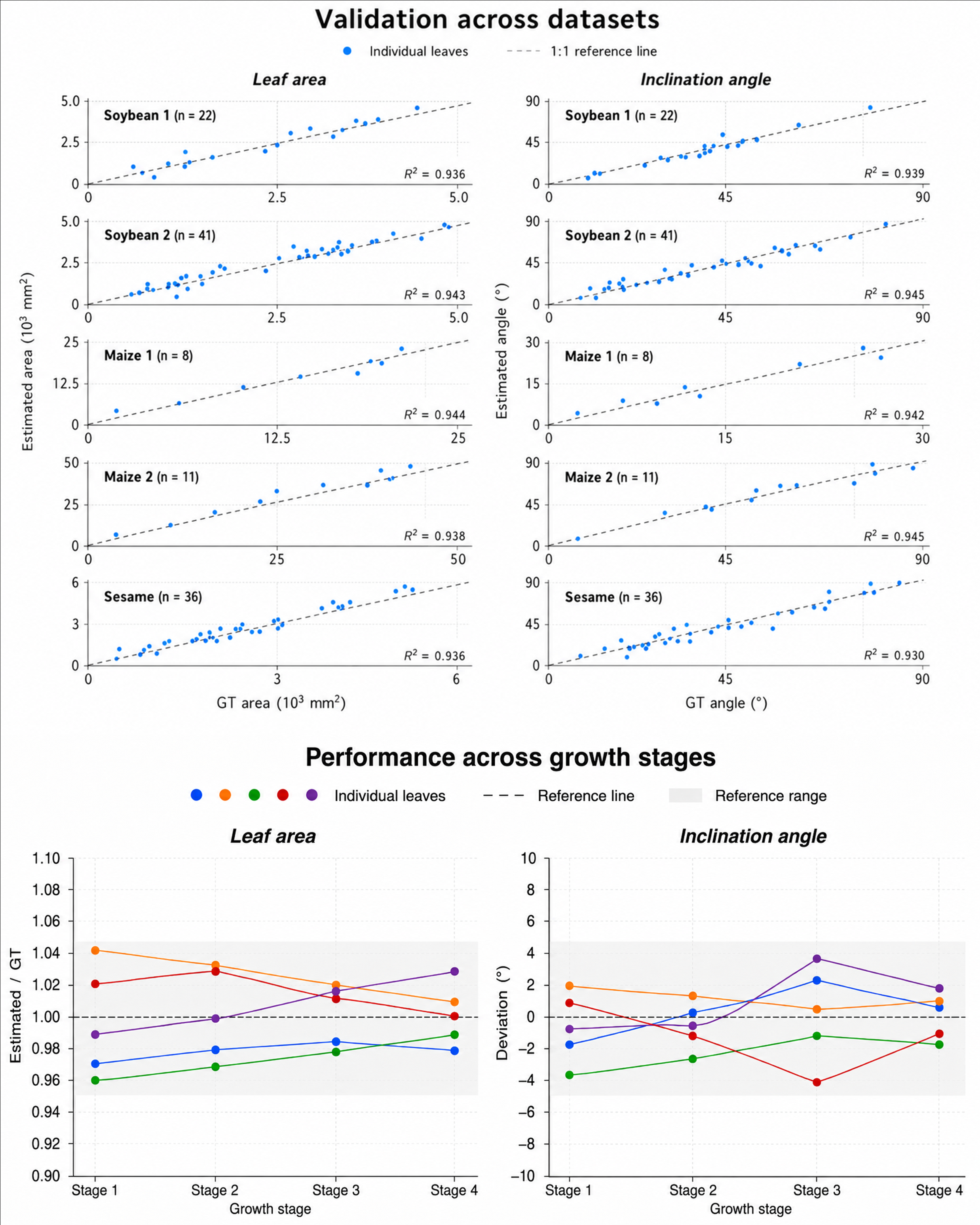
在5种成熟植物(大豆、芝麻、玉米)上的定量评估显示:叶面积估计值与真值的比值在0.9514-1.0629之间,叶倾角平均绝对误差约2.04°。在36个玉米幼苗时序序列上也表现出稳定的跨阶段一致性。
消融实验
| 稀疏初始化 | 密集模型 | 新视角增强 | PSNR | 叶面积 R² | 叶倾角 R² | 总时间 |
|---|---|---|---|---|---|---|
| COLMAP | 3DGS | × | 27.98 | 0.898 | 0.910 | 37.3 min |
| COLMAP | Mip | ✓ | 28.95 | 0.924 | 0.934 | 39.1 min |
| π³ | Mip | ✓ | 29.86 | 0.941 | 0.946 | 32.6 min |
三个设计选择(3DFM初始化、Mip-Splatting密集化、新视角增强)均有贡献,其中稀疏初始化贡献最大。π³+Mip+新视角增强的完整配置在PSNR(29.86)、叶面积R²(0.941)和叶倾角R²(0.946)上均达到最佳,同时总时间从39.1分钟降至32.6分钟。
个人见解 Insights
这篇论文将3D基础模型(3DFMs)引入植物表型领域,是一个非常及时且有实际价值的工作。以下是几点观察:
- 优势:系统性验证了3DFMs在植物场景的可行性,不是简单地应用大模型,而是设计了完整的从重建到测量的流水线。跨作物(10种)+多环境(室内/室外/田间)+多生长阶段的验证覆盖面广。
- 优势:提供了开源的跨作物3D表型数据集,包含器官级标注和手动表型真值,对社区贡献显著。
- 不足:当前验证主要在单株、近距离、闭环采集条件下,密集冠层、田间大规模场景的适用性尚未充分验证。
- 不足:度量尺度恢复依赖花盆几何作为场景内参考,在无花盆的田间部署中需要替代方案(平台位姿、行间距等)。
- 启发:3DFM的前馈式几何推理范式正在从通用3D重建向垂直领域渗透。植物表型是一个典型受益场景——薄叶片、重复纹理、自遮挡正是传统SfM的痛点,而3DFM的全局先验恰好能克服这些困难。
- 启发:2D-to-3D语义迁移策略(多视角2D分割→反投影→融合)相比直接3D分割的巨大优势(mIoU提升40%+),暗示在稀疏/不规则3D点云上,利用成熟的2D视觉模型可能是更务实的路径。
参考资料 References
查看原文 View Original Paper
arXiv: 2607.01753
下载 PDF Download PDF
arXiv PDF